منبع سیگنال EEG
منبع سیگنال EEG: نقش پتانسیلهای پسسیناپسی در تولید امواج مغزی
چکیده
الکتروانسفالوگرافی (Electroencephalography یا EEG) یکی از مهمترین روشهای غیرتهاجمی برای مطالعه فعالیت الکتریکی مغز است که در علوم اعصاب، نورولوژی، روانپزشکی و علوم شناختی کاربرد گستردهای دارد. با وجود استفاده فراوان از EEG در تحقیقات و تشخیصهای بالینی، هنوز یکی از پرسشهای بنیادی در میان دانشجویان و پژوهشگران این است که **سیگنال واقعی EEG از چه منبعی در مغز تولید میشود**. برخلاف تصور رایج، سیگنال EEG حاصل شلیک پتانسیلهای عمل نورونها نیست، بلکه عمدتاً بازتابی از **پتانسیلهای پسسیناپسی (Postsynaptic Potentials)** در دندریتهای نورونهای هرمی قشر مغز است. این مقاله با رویکردی علمی و آموزشی، به بررسی دقیق فیزیولوژی پتانسیلهای پسسیناپسی، نحوه ایجاد میدانهای الکتریکی در بافت قشری، نقش همزمانی نورونی و مکانیسم انتقال این میدانها تا سطح جمجمه میپردازد. هدف این مقاله ارائه یک چارچوب مفهومی روشن برای درک منبع سیگنال EEG و بهبود تفسیر علمی دادههای EEG و qEEG است.
مقدمه
مغز انسان شبکهای فوقالعاده پیچیده متشکل از میلیاردها نورون است که از طریق سیناپسها با یکدیگر ارتباط برقرار میکنند. این ارتباطات الکتروشیمیایی اساس تمام فرآیندهای شناختی، حسی و حرکتی را تشکیل میدهند. یکی از مهمترین ابزارهایی که برای مطالعه فعالیت الکتریکی مغز توسعه یافته است، **الکتروانسفالوگرافی (EEG)** است.
EEG برای اولین بار در سال ۱۹۲۴ توسط **هانس برگر (Hans Berger)** معرفی شد. این روش امکان ثبت نوسانات الکتریکی مغز را از طریق الکترودهایی که روی پوست سر قرار میگیرند فراهم میکند. امواجی که در EEG مشاهده میشوند شامل ریتمهایی مانند دلتا، تتا، آلفا، بتا و گاما هستند که هر یک با حالتهای مختلف عملکرد مغز مرتبطاند.
با این حال، سؤال کلیدی این است:
**سیگنالهایی که توسط EEG ثبت میشوند دقیقاً از کدام فعالیت عصبی ناشی میشوند؟**
تحقیقات گسترده در علوم اعصاب نشان دادهاند که منبع اصلی سیگنال EEG، **پتانسیلهای پسسیناپسی همزمان در دندریتهای نورونهای هرمی قشر مغز** است. این پتانسیلها در اثر جریان یونها از طریق کانالهای غشایی در پاسخ به انتقالدهندههای عصبی ایجاد میشوند.
درک این مکانیسم برای تفسیر صحیح EEG، تحلیل دادههای qEEG و طراحی مطالعات علوم اعصاب اهمیت بنیادی دارد.
پتانسیلهای پسسیناپسی چیستند؟
برای درک منبع سیگنال EEG ابتدا باید مفهوم **پتانسیل پسسیناپسی (Postsynaptic Potential)** را بررسی کنیم.
در سیستم عصبی، ارتباط بین نورونها از طریق ساختاری به نام **سیناپس** انجام میشود. در یک سیناپس شیمیایی، نورون پیشسیناپسی هنگام فعال شدن، انتقالدهندههای عصبی را به فضای سیناپسی آزاد میکند. این مولکولها به گیرندههای موجود روی غشای نورون پسسیناپسی متصل میشوند و موجب تغییر در نفوذپذیری یونی غشا میگردند.
این تغییر نفوذپذیری باعث ایجاد یک تغییر ولتاژ در غشای نورون پسسیناپسی میشود که به آن **پتانسیل پسسیناپسی** گفته میشود.
پتانسیلهای پسسیناپسی به دو نوع اصلی تقسیم میشوند:
a- پتانسیل پسسیناپسی تحریکی (EPSP)
b- پتانسیل پسسیناپسی مهاری (IPSP)
این دو نوع پتانسیل اساس پردازش اطلاعات در مغز را تشکیل میدهند.
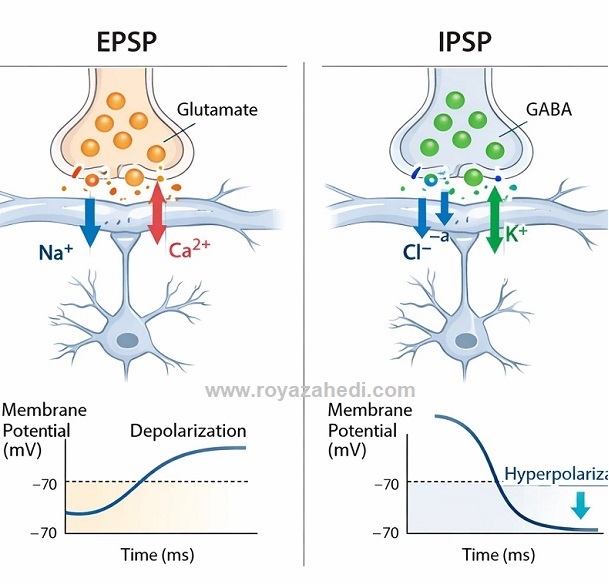
پنل سمت چپ با برچسب EPSP:
یک سیناپس تحریکی روی دندریت یک نورون هرمی نشان داده شده است. وزیکولهای گلوتامات آزاد میشوند و یونهای سدیم (Na⁺) و کلسیم (Ca²⁺) به داخل دندریت وارد میشوند. نمودار تغییرات ولتاژ غشا نیز نشاندهنده دپولاریزاسیون و افزایش ولتاژ به سمت بالا است.
پنل سمت راست با برچسب IPSP:
یک سیناپس مهاری با وزیکولهای GABA نمایش داده شده است. یونهای کلر (Cl⁻) وارد سلول میشوند و یونهای پتاسیم (K⁺) از سلول خارج میشوند. نمودار ولتاژ غشا هایپرپولاریزاسیون و کاهش ولتاژ به سمت پایین را نشان میدهد.
در تصویر یک نورون هرمی سادهشده با دندریتها، شکاف سیناپسی، فلشهای نشاندهنده حرکت یونها و نمودارهای کوچک ولتاژ-زمان نیز وجود دارد.
پتانسیل پسسیناپسی تحریکی (EPSP)
پتانسیل پسسیناپسی تحریکی زمانی ایجاد میشود که فعال شدن سیناپس باعث **دپولاریزاسیون غشای نورون** شود. در این حالت، ولتاژ غشا به سمت مقادیر مثبتتر حرکت میکند و نورون به آستانه شلیک نزدیکتر میشود.
مهمترین انتقالدهنده عصبی تحریکی در مغز **گلوتامات (Glutamate)** است. این انتقالدهنده به گیرندههایی مانند AMPA و NMDA متصل میشود و موجب باز شدن کانالهای یونی سدیم و کلسیم میگردد. ورود یونهای مثبت به داخل نورون باعث کاهش اختلاف پتانسیل غشایی و ایجاد EPSP میشود.
EPSPها معمولاً دامنهای در حد چند میلیولت دارند و مدت زمان آنها بین ۱۰ تا ۱۰۰ میلیثانیه است. این زمان نسبتاً طولانی باعث میشود که EPSPها بتوانند با یکدیگر جمع شوند و اثر قابل توجهی در فعالیت نورون ایجاد کنند.
پتانسیل پسسیناپسی مهاری (IPSP)
در مقابل EPSP، پتانسیل پسسیناپسی مهاری باعث **هایپرپولاریزاسیون غشای نورون** میشود. در این حالت، ولتاژ غشا منفیتر میشود و احتمال شلیک نورون کاهش مییابد.
مهمترین انتقالدهنده عصبی مهاری در مغز **GABA (گاما آمینوبوتیریک اسید)** است. اتصال GABA به گیرندههای GABA-A معمولاً باعث باز شدن کانالهای کلر میشود و ورود یونهای کلر به داخل سلول غشا را منفیتر میکند. در برخی موارد نیز خروج یونهای پتاسیم باعث مهار نورون میشود.
IPSPها نیز مشابه EPSP دارای مدت زمان نسبتاً طولانی هستند و میتوانند با سایر پتانسیلها جمع شوند.
چرا EEG پتانسیلهای عمل را ثبت نمیکند؟
یکی از اشتباهات رایج در درک EEG این است که تصور میشود EEG مستقیماً شلیک نورونها یا **پتانسیلهای عمل (Action Potentials)** را ثبت میکند. در واقع چنین نیست.
پتانسیلهای عمل بسیار سریع هستند و مدت زمان آنها حدود یک میلیثانیه است. علاوه بر این، این پتانسیلها در نورونهای مختلف به صورت نامنظم و ناهمزمان رخ میدهند. به همین دلیل، میدانهای الکتریکی تولید شده توسط آنها به سرعت یکدیگر را خنثی میکنند و به سطح جمجمه نمیرسند.
در مقابل، پتانسیلهای پسسیناپسی بسیار کندتر هستند و میتوانند در هزاران نورون به طور همزمان رخ دهند. این ویژگی باعث میشود میدانهای الکتریکی آنها با یکدیگر جمع شوند و سیگنالی قابل ثبت توسط EEG ایجاد کنند.
نقش نورونهای هرمی در تولید سیگنال EEG
یکی از مهمترین کشفیات در علوم اعصاب این است که **نورونهای هرمی قشر مغز منبع اصلی سیگنال EEG هستند**.
نورونهای هرمی ویژگیهایی دارند که آنها را برای تولید میدانهای الکتریکی قابل ثبت مناسب میکند:
اول اینکه این نورونها به صورت **منظم و عمود بر سطح قشر مغز** قرار گرفتهاند. این جهتگیری منظم باعث میشود میدانهای الکتریکی آنها همدیگر را تقویت کنند.
دوم اینکه نورونهای هرمی دارای **دندریت رأسی بلند** هستند که از لایههای عمیق قشر به سمت سطح قشر امتداد مییابد. بیشتر سیناپسهای تحریکی روی همین دندریتها قرار دارند.
سوم اینکه این نورونها تعداد بسیار زیادی دارند و بخش عمده نورونهای قشری را تشکیل میدهند. در نتیجه، فعالیت همزمان آنها میتواند میدانهای الکتریکی بزرگی ایجاد کند.
ایجاد دیپل الکتریکی در نورون
وقتی یک EPSP در دندریت نورون هرمی رخ میدهد، جریان یونی به داخل سلول وارد میشود. این ورود یونهای مثبت باعث میشود محل سیناپس نسبت به محیط اطراف بار منفیتری داشته باشد.
به این ناحیه **sink** گفته میشود. در مقابل، جریان الکتریکی از بخش دیگری از نورون خارج میشود که به آن **source** گفته میشود.
این توزیع بار الکتریکی در طول نورون یک ساختار **دیپل (Dipole)** ایجاد میکند. هر نورون فعال در واقع یک دیپل کوچک تولید میکند.
وقتی هزاران یا میلیونها نورون هرمی به طور همزمان فعال شوند، دیپلهای آنها با یکدیگر جمع میشوند و میدان الکتریکی بزرگی ایجاد میکنند که میتواند از طریق بافت مغز، مایع مغزی–نخاعی، جمجمه و پوست سر عبور کند و توسط الکترودهای EEG ثبت شود.
اهمیت همزمانی نورونی
برای اینکه سیگنال EEG قابل اندازهگیری باشد، لازم است تعداد زیادی نورون به صورت همزمان فعال شوند. تخمین زده میشود که حداقل **دهها هزار تا یک میلیون نورون** باید به طور همزمان در یک ناحیه فعال شوند تا سیگنال قابل ثبت ایجاد شود.
این پدیده به عنوان **همزمانی نورونی (Neural Synchrony)** شناخته میشود.
همزمانی نورونی نقش مهمی در ایجاد ریتمهای مغزی دارد. برای مثال، امواج آلفا که در حالت استراحت با چشمهای بسته دیده میشوند، نتیجه فعالیت هماهنگ شبکههای تالاموکورتیکال هستند.
هرچه همزمانی نورونها بیشتر باشد، دامنه سیگنال EEG نیز بزرگتر خواهد بود.
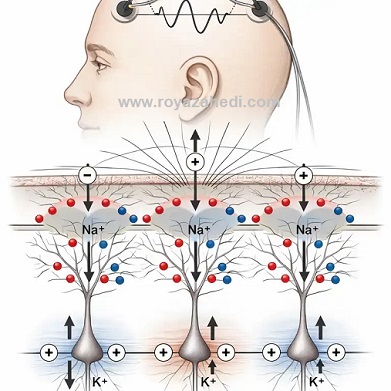
تصویر منبع سیگنالهای EEG و نقش پتانسیلهای پسسیناپسی** را نشان میدهد.
در تصویر، **نورونهای هرمی قشر مغز** به صورت **همراستا و عمود بر سطح قشر** نمایش داده شدهاند و **دندریتهای رأسی (apical dendrites)** بهوضوح قابل مشاهده هستند.
**ورودیهای سیناپسی تحریکی (EPSP)** و **مهاری (IPSP)** روی دندریتها با **کدگذاری رنگی** مشخص شدهاند (قرمز برای EPSP و آبی برای IPSP).
جریانهای یونی بهگونهای ترسیم شدهاند که **ناحیههای منبع (source) و چاهک (sink)** را ایجاد کرده و منجر به **تشکیل دیپلهای الکتریکی** میشوند.
چندین نورون هرمی که بهصورت **همزمان فعال شدهاند** نشان داده میشوند که اثر آنها با هم **جمع (summation)** شده و یک **میدان الکتریکی بزرگ** ایجاد میکند.
در بخش بالایی تصویر، **الکترودهای EEG روی پوست سر** نمایش داده شدهاند که این پتانسیلهای جمعشده را ثبت میکنند.
انتقال میدان الکتریکی از قشر مغز به اسکالپ
میدان الکتریکی ایجاد شده توسط نورونهای قشری باید از چند لایه عبور کند تا به الکترودهای EEG برسد. این لایهها شامل موارد زیر هستند:
a- بافت مغز
b- مایع مغزی–نخاعی
c- جمجمه
d- پوست سر
هر یک از این لایهها مقاومت الکتریکی خاصی دارند و باعث کاهش شدت سیگنال میشوند. به همین دلیل دامنه سیگنال EEG در سطح اسکالپ معمولاً در حد **چند میکروولت** است.
این کاهش شدت همچنین باعث میشود EEG وضوح مکانی محدودی داشته باشد.
ارتباط پتانسیلهای پسسیناپسی با ریتمهای EEG
ریتمهای مختلف EEG بازتاب الگوهای متفاوتی از فعالیت شبکههای عصبی هستند. این ریتمها در واقع حاصل تغییرات در همزمانی EPSPها و IPSPها در جمعیت بزرگی از نورونهای هرمی هستند.
برای مثال:
a- امواج دلتا معمولاً در خواب عمیق دیده میشوند و نشاندهنده فعالیت هماهنگ گسترده در قشر مغز هستند.
b- امواج تتا با فرآیندهای حافظه و یادگیری مرتبطاند.
c- امواج آلفا در حالت استراحت و بستن چشمها افزایش مییابند.
d- امواج بتا و گاما با پردازش شناختی فعال و توجه مرتبط هستند.
اهمیت درک منبع سیگنال EEG در پژوهش و بالین
درک اینکه سیگنال EEG حاصل پتانسیلهای پسسیناپسی است، پیامدهای مهمی برای تفسیر دادههای EEG دارد.
اول اینکه این موضوع نشان میدهد EEG بیشتر بازتاب **فعالیت دندریتها و پردازش سیناپسی** است، نه صرفاً شلیک نورونها.
دوم اینکه تغییرات در ریتمهای EEG میتواند نشاندهنده تغییر در **تعادل بین تحریک و مهار سیناپسی** باشد.
سوم اینکه در تحلیلهای پیشرفته مانند qEEG، پارامترهایی مانند توان طیفی، همدوسی و فاز در واقع شاخصهایی از الگوهای همزمانی در شبکههای نورونی هستند.
نتیجهگیری
الکتروانسفالوگرافی یکی از ارزشمندترین ابزارها برای مطالعه عملکرد مغز است، اما درک صحیح آن نیازمند شناخت دقیق منبع سیگنالهای ثبتشده است. شواهد گسترده در علوم اعصاب نشان میدهد که سیگنال EEG عمدتاً حاصل **پتانسیلهای پسسیناپسی در دندریتهای نورونهای هرمی قشر مغز** است.
این پتانسیلها در نتیجه جریان یونها در پاسخ به فعالیت سیناپسی ایجاد میشوند و هنگامی که تعداد زیادی از نورونها به صورت همزمان فعال شوند، میدانهای الکتریکی آنها با یکدیگر جمع شده و سیگنال قابل ثبت توسط EEG را ایجاد میکنند.
درک این مکانیسم پایهای، نه تنها برای دانشجویان علوم اعصاب بلکه برای پژوهشگران و متخصصانی که با EEG و qEEG کار میکنند ضروری است، زیرا این دانش چارچوبی علمی برای تفسیر دقیقتر دادههای مغزی فراهم میکند.